Research Project On The Role Of The Innate Immunity In The Pathogenesis Of Pediatric Malignancies And Autoimmune Disease

Project location: Italy, Rome
Project start date: July 2009 -
Project end date: January 2010
Project number: 2009-41
Beneficiary: Arcobaleno ONLUS
The LAK phenomenon is identified by the ability of cytokine activated peripheral blood lymphocytes (PBL) to generate cytotoxic cells capable of eliminating solid tumor cells and hematopoietic tumor cell lines, without previous immunization or MHC restriction. This phenomenon is mediated by the NK cells and to a lesser extent by NK-T cells.
Translational studies have attempted to utilize these cells in a clinical setting. Large numbers of PBMC were stimulated in vitro with high doses of interleukin-2 (IL-2) and adoptively transferred in patients affected by haematological and non haematological malignancies including leukemias, renal cell carcinoma and melanoma.
Unfortunetely, only a small subset of these patients benefited of such a immuno therapy. Possible explanations for the resistance of cancer cells to LAK cells could be explained by the missing self hypothesis. HLA-class I positive cancer cells would bind killer inhibitory receptors (KIRs) on autologous LAK cells, leading to the neutralization of autologus NK cell cytotoxicity. Alternatively, host cancer cells may utilize defensive mechanisms of escape from autologous NK cell recognition. However, recent studies have shown that HLA-class I positive leukemia cells expressing MICA/B antigen can be killed by HLA-matched or autologous NK cells. Therefore, the mechanism of malignant cell resistance to NK cell cytotoxicity may involve multiple pathways.
Based on our laboratory experience and literature data, in this study he has hypothesized that the failure of HLA-matched or autologous NK cells to control leukemia cells involves e mechanism of elimination of NK cells by leukemia cells which leads to the
mutual cell elimination.
The aim of this study intended to verify the following hypothesis
1. Leukemia and cancer cells avoid NK cell recognition by triggering NK cell elimination though an apoptotic/necrotic mechanism
2. The mechanism of NK cells elimination by malignant cells involves the Fc gamma receptor III (FcγRIII) expressed on NK cells.
3. The interaction between diabetic beta cells and NK cells results in beta cell elimination without affecting NK cell survival and therefore triggering NK cell autoimmunity toward MICA/B+ beta cells.
Accomplishment
Results and Discussion
Effects of myeloid leukemia cells on IL-2 stimulated, allogeneic NK cells.
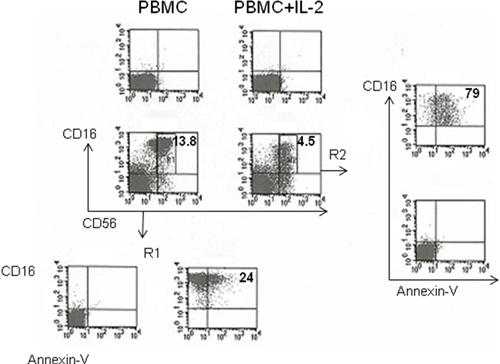
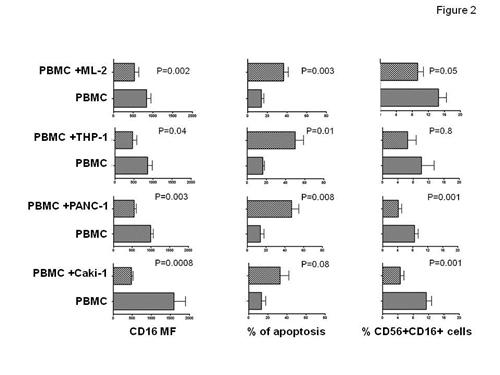
In this study he has first assessed the effect of the tumor cell line HCT-116 on IL-2 stimulated, cytotoxic NK cells. Figure 1 shows that HCT116 stormed NK cells by increasing elimination, apoptosis and decreasing the surface expression of CD56+CD16+ cells, suggesting that cancer cells deploy mechanisms of evasion from NK cell immunesurveillance. He next evaluated extent of cancer cells dependent NK cell perturbation utilizing either leukemia cell lines or epithelial cancer cell lines. Figure 2 shows that a differential capability of inducing NK cell perturbation between leukemia and cancer epithelial cells. Interestingly, he found that there was e differential extent of malignant cell induced NK cell perturbation. ML-2 and THP-1 preferentially induced CD56+CD16+ NK cell apoptosis and CD16 down-regulation while PANC-1 and Caki-1 cells reduced CD56+CD16+ NK cell frequency, apoptosis and CD16 down-regulation.
Specificity of tumor cell induced NK cell perturbation
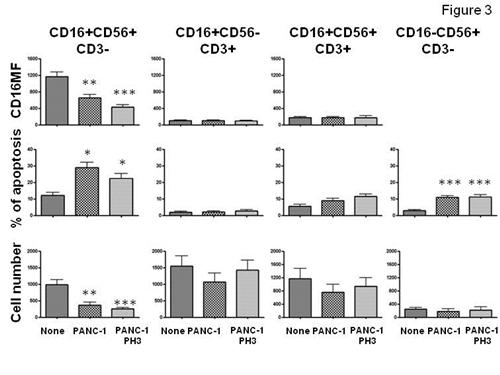
In order to evaluate the specificity of cancer cell induced NK cell perturbation. He analyzed the effects of PANC-1 cells on Lymphokine (IL-2) activated killer cells (LAK). Figure 3 shows that PANC-1 cells preferentially perturbed the most, cytotoxic, NK cell subset (CD16+CD56+CD3- cells) and a to lesser extent the least (CD16-CD56+CD3- cells) cytotoxic NK cell subset. In contrast, CD16+ and CD16- NK-T cells were not significantly affected by PANC-1 cells. Since NK cell cytotoxicity is profoundly influenced by the surface level of HLA-class-I antigen, he asked whether or not HLA-class I antigens would affect cancer cell dependent NK cell perturbation. As shown in Figure 3, CD16MF, absolute NK and NK-T cell number and NK-NK-T cell apoptosis of PBMC cultured in the presence of PANC-1 cells with or without HA-class I cell surface expression did not show significantly different results suggesting that PANC-1 induced NK cell perturbation does not require HLA-class I antigen expression. Similar results were observed when other tumor cell lines including ML-2 or THP-1 were utilized (data not shown). To further evaluate the specificity of NK cell perturbation he tested the effect of mouse leukemia cell line, RAW.264.7 (H2d) on IL-2 stimulated C57BL/6J (B16, H2b) splenocytes.
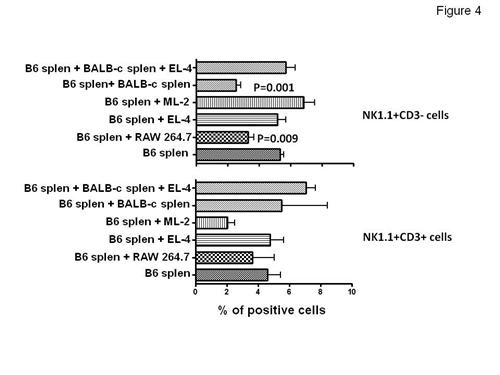
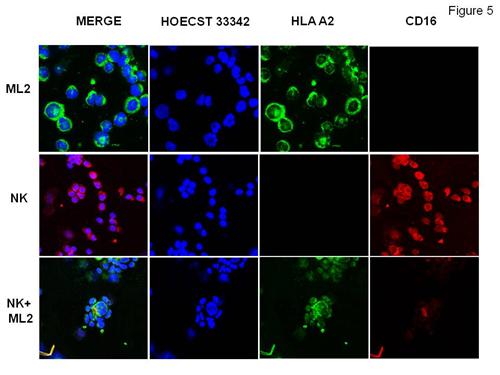
Figure 4, left panel, shows that RAW264.7 cells induced B16, NK1.1+CD3-NK cells elimination while syngeneic EL4 cells did not. Furthermore, BALB/c (H2d) allogeneic, splenocytes triggered B16 mouse NK cell elimination. Interestingly, pre-incubation of BALB/c splenocytes with EL4 cells at 4:1 ratio protected B16 mouse NK cells from BALB/c splenocytes suggesting that allogeneic EL4 neutralized the effects of BALB/c splenocytes on B16 NK cells by neutralizing BALB/c NK cells. These data strongly support the idea that mouse NK cells cultured with allogeneic tumor cells but not with normal, allogeneic, splenocytes undergo alloperturbation phenomenon which is specifically triggered by tumor cells. This phenomenon was restricted to NK cells and did not involve NK-T cells (Fig.4 right panel). To evaluate the relationship of CD16 molecules on the alloperturbation phenomenon, he cultured ML2 HLA- A2 + antigen with magnetically sorted HLA-A2 negative NK cells. Then the path of CD16 was analyzed during effector/target cell conjugation by confocal microscopy. He found a strong polarization and co-localization of CD16 with HLA-A2 in the immunological synapses raised by NK-ML2 cell contact (Fig.5). These data strongly suggest that ML-2 express an unknown CD16 receptor
In summary, to date, we present evidence showing that a wide range of cell lines induce (i) specifically perturbation (CD16 down-regulation, NK cell elimination, and NK cell apoptosis) of allogeneic NK cells; (ii) this phenomenon is also identified in mouse NK cells but not in NK-T or T cells; (iii) CD16 polarizes at the immunological synapses formed between NK/ML-2 or THP-1 conjugation suggesting that tumor cells bear a CD16 specific receptor.
Publications
1. Tumor infiltration by FcγRIII (CD16)+ myeloid cells is associated with improved survival in patients with colorectal carcinoma. Sconocchia G, Zlobec I, Lugli A, Calabrese D, Iezzi G, Karamitopoulou E, Patsouris ES, Peros G, Horcic M, Tornillo L, Zuber M, Droeser R, Muraro MG, Mengus C, Oertli D, Ferrone S, Terracciano L, Spagnoli GC.Int J Cancer. 2011 Jun 1;128(11):2663-72. doi: 10.1002/ijc.25609. Epub 2010 Oct 13
2. Leukemia and epithelial cancer cells triggered NK cell storming : The NK cell perturbation phenomenon
Arriga R, Caratelli S, Basello K, Coppola A, Capuani B, Lauro D, Donadel G, Ferrelli F, Spagnoli GC, Sconocchia G. 2011 Submitted
| FIGURE | CAPTION |
| Figure 1 |
HCT-116 induced NK cell elimination, apoptosis and CD16 down-regulation PBMC were cultured for 3 days in the presence of IL-2. Cells were harvested and cultured overnight in the presence or absence of HCT-116 as indicated. R1 and R2 are electronic gates posted on CD56+CD16+ cytotoxic cells. The numbers shown in the quadrant plots are the percentages of positive cells. This figure represents one of 4 representative experiments performed with similar results.
|
| Figure 2 |
Effects of leukemia and epithelial cancer cell lines on CD56+CD16+ NK cells. IL-2 stimulated PBMC were cultured in the presence or absence of ML-2 (data from 6 experiments), THP-1 (data from 4 different experiments), PANC-1 (data from 4 different experiments), Caki-1 (data from 10 different experiments)
|
| Figure 3 | Cancer cell induced NK cell perturbation preferentially involves CD16+ and CD16- NK cell subsets and does not require cell surface HLA-class I antigen expression. IL-2 stimulated PBMC were cultured in the presence or absence of PANC-1 cells untreated or treated at PH3 for 2 minutes. After a 5-hour cell culture, NK and NK-T cell subsets analysis was performed by flow cytometry Upper left panel: ** = none vs PANC-1 (P=0.002); *** = none vs PANC-1PH3 ( P=0.001) ; Middle left panel: **= none vs PANC-1 (P=0.002) and none vs PANC-1PH3 (P=0.001); Lower left panel: *= none vs PANC-1 (P=0.03) and none vs PANC-1PH3 (P=0.03) , Lower right panel: *** = none vs PANC-1 (P=0.0001) and none vs PANC-1PH3 (P=0.0006). No statistically significant differences between PANC-1 and PANC-1PH3 conditions were identified. Data obtained from 6 separated experiments
|
| Figure 4 |
Cancer cell induced NK cell alloperturbation involves mouse NK cells and requires allogeneic cancer cells. B16 splenocytes were stimulated for 3 days with IL-2 and cultured in the presence or absence of the indicated cells. Left panel % of positive cells; right panel % of annexin-V positive cells.
|
| Figure 5 |
CD16 molecule polarization and colocalization at the immunological synapses during NK/ML-2 leukemia cell conjugation.
|